|
|
|


概日リズムを持つ原核生物
概日時計を有する生物は数多く報告されているが,我々のグループは現在のところシアノバクテリアを選択している (図1)。シアノバクテリアは生命進化の初期に水を分解する光合成機能を獲得し、その後の多様な生命進化を可能した生物である。この生物は核をもたない原核生物であるが、光合成に依存した生活を営むため時計機能を持っており、最も単純なモデル生物として好都合であった.
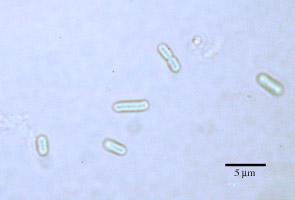
図1:シアノバクテリアの顕微鏡写真
シアノバクテリアの生物時計をみる
シアノバクテリアの示す様々な生命現象のうち,恒明条件化で概日リズムを示すことが知られているのは光合成活性,窒素固定活性などである.こうした生理学的な観察をする方法もあるが,我々のグループはある概日リズムを刻むことがすでにわかっている遺伝子の発現を生物発光を利用してリアルタイムに観察している.シアノバクテリア自身の生物時計はシアノバクテリアのなかには遺伝子操作の容易な種(Synechococcus PCC 7942)がある。これに概日時計に制御されるよう仕組んだ生物発光酵素の遺伝子を移入し、生物発光を計ることで概日時計の動きを容易にモニターすることができる.生物発光はもちろん連続して何日も計測しなければならないが,自動的に長時間の計測をしてくれる近藤先生お手製の測定装置を用いて簡単に測ることができる(図2).(噂によればこの装置は時間生物学業界ではコンドートロンと呼ばれているらしい)

図2:生物発光測定装置
シアノバクテリアの時計関連遺伝子
近藤研の初期の研究は,シアノバクテリアの遺伝子の中から概日時計と関わっている遺伝子(時計関連遺伝子とよぶ)を探すことであった.先ほど述べた生物発光リズムは寒天上に成育した多数のコロニーでも観測することができるので、容易にリズム突然変異を分離することができた。これらの変異体では概日時計に不可欠な遺伝子が異常をおこしていることが考えられるため、石浦博士(現遺伝子実験施設)らとともに、この遺伝子を突き止めようとした。まず正常なゲノムを小さな断片に分解し、1つずつ突然変異体の細胞に導入する。もし突然変異を起こした遺伝子に相当する正常な遺伝子含むゲノム断片が入れば、リズムは元に戻る。これが見つかればそのなかに時計遺伝子が存在するはずである。こうして3つの遺伝子からなる時計遺伝子群kaiABC(回転にちなんでkaiと命名した)が見つけられた.またその後も突然変異体の観察から時計遺伝子を探す試みは続けられた.現在までに我々のグループは時計関連遺伝子としてkaiABCに加え、sasA, rpaA, labAを報告しており、他のグループからもいくつかの遺伝子が報告されている.転写翻訳ネガティブフィードバック説とKaiC
生物時計はどのような分子的なメカニズムにより生じるのだろう?多くの生物において支持されてきた説は"転写翻訳ネガティブフィードバック説"である(図3).
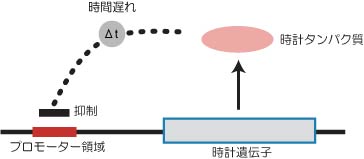
図3:転写翻訳ネガティブフィードバック説
これは,ある遺伝子から発現したタンパク質がある時間経過した後に,その遺伝子自身の発現を抑えるというモデルである.ある時計遺伝子がまず発現しタンパク質を生産したとする.時計タンパク質は少しの時間をおいた後,今度は自身の発現を抑制するので遺伝子発現はストップする.そのうち時計タンパク質は分解し量が徐々に減ってくる.そうするとまた時計遺伝子の発現が開始され,最初の状態にもどる.このような仕組みにより、振動がおこる。他の生物ですでに報告されていたと時計関連遺伝子はよくこのモデルに適合した.シアノバクテリアの場合はkaiCがこのモデルに適合し,kaiCを中心とした転写翻訳ネガティブフィードバックループモデルが振動を生み出すと考えられた(図4).
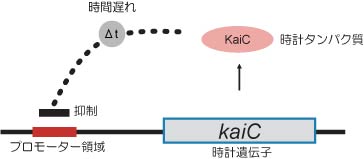
図4:KaiCによるネガティブフィードバックループ
KaiCリン酸化リズムの発見と転写翻訳ネガティブフィードバック説への疑い
KaiCの遺伝子発現制御とは別に,KaiC自身の生化学的性質への研究も同時に進められた.その過程でKaiCは細胞内においてATPを加水分解し自身をリン酸化することが明らかになった.このリン酸化反応とKaiA, KaiBタンパク質は関係しており,kaiA, KaiBの新たな役割も浮かび上がった.また重要なことは細胞内においてリン酸化されたKaiCの割合が24時間周期で変動している,すなわち概日リズムを刻んでいることである.
この"KaiCのリン酸化のリズム"に対して,2004年頃,不思議な現象が観察された.シアノバクテリアは恒暗条件下でほとんどの遺伝子発現がストップすることがわかっていた.その条件下でもあいかわらずKaiCのリン酸化のリズムは継続していることがわかった.すなわちこのリン酸化リズムは転写翻訳ネガティブフィードバックとは関わらずに存在していることが示唆される.この結果からシアノバクテリアにおいては転写翻訳ネガティブフィードバックモデルではなく,KaiCのリン酸化リズムが概日リズムを生み出す装置そのものではないかとの推測された.
Kaiタンパク質による概日時計の試験管内再構成
KaiCタンパク質の細胞内での不思議な振る舞いから間もない2004年秋ごろ、我々のグループでついにリン酸化リズムそのものが試験管内で再構成された.しかもその系はKaiCのほかに必要なものはKaiAとKaiBタンパク質,それとATPのみという, 極めてシンプルな系であった. これは,KaiCのリン酸化リズムが自律的に振動する能力が存在することを指し示すものであり,KaiCのリン酸化リズムがシアノバクテリアの概日時計の中心であるということを示す結果である.細胞内では,KaiCのリン酸化のレベルを検知して各種の遺伝子発現制御を行っているというモデルが今のところ細胞の概日リズムを生み出すメカニズムとして有力である(図5).
試験管内でのKaiCのリン酸化リズムは概日時計の3つの条件(約24時間周期、位相同調性、周期の温度補償性、「概日時計とは?」を参照)を満たす完璧な概日リズムである。一方この系はBZ反応のような,一定環境下で自律的に駆動する化学振動子としての側面も持っている.生物時計だけの問題ではなく,純粋な化学,物理学の問題としてとらえても解くべき謎は多い.我々のグループだけではなく,多くのグループがこの系に着目し研究を進めている.
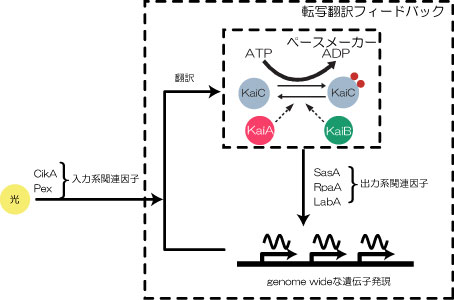
図5:KaiCリン酸化リズムが細胞全体の概日時計を駆動する
KaiCのATP分解活性〜KaiCの中に時計がある!〜
シアノバクテリアの概日時計の主役であるKaiCは2つのATP分解酵素(ATPase)がつながって構成されている。ATPは分解するとエネルギーを発生する、ほとんどの生命活動のエネルギー源になる分子である。生命はATPaseを使ってATPを分解する事で生命活動のためのエネルギーを得ている。2007年から我々のグループはKaiCのATPase活性に着目し、研究を行ってきた。これまでの我々の研究により、次の事が明らかになった(図6)。
① KaiCのATP活性は1分子のKaiCで1日にわずか10から15個のATPを分解するのみ(多くの酵素活性の1万分の1以下)と非常に低い。
② しかし、KaiCのATPase活性は極めて安定で温度の影響をうけない。
③ さらに周期変異体のKaiCで調べると、KaiCのATPase活性は概日時計の速度に比例する。
④ ①から③の性質はKaiCだけで見られ、KaiAやKaiBを必要としない。
これらの結果は温度補償性と周期という概日時計の特性が、KaiAやKaiBとの相互作用を介さず、KaiCのみのATPase反応で決まることを示している。すなわち、KaiCのATPase反応はシアノバクテリアの概日時計のペースメーカーであると考えられる。
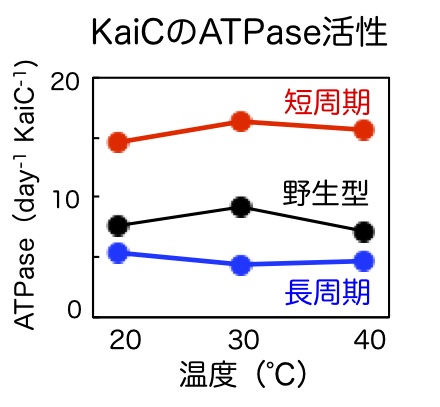
図6:KaiCのATPase活性は温度の影響を受けず、時計の速度に比例する。
現在の我々のグループの研究
KaiCのATPase反応が温度補償性と周期を決める仕組み、そしてATPase反応が振動を発生させる仕組みを解明することを目指している。現在、KaiCのATPase活性が温度補償される仕組みとして、KaiCのATP分解エネルギーがKaiC自身の構造にバネのようなひずみを生じることによって自身のATPase活性を抑制するモデル(分子内フィードバック)を1つの可能性として想定している(図7)。KaiCは2つのATP分解酵素がつながって構成されているが、一方(CIドメイン)はATPase活性の大部分、もう一方(CIIドメイン)はリン酸化サイクルという異なる機能を分担する。2つのATP分解酵素がどのように関わりあって、どの温度でも約24時間周期の安定なリズムを生じるのか、総力戦でこの分子機械の理解に挑んでいるところである.
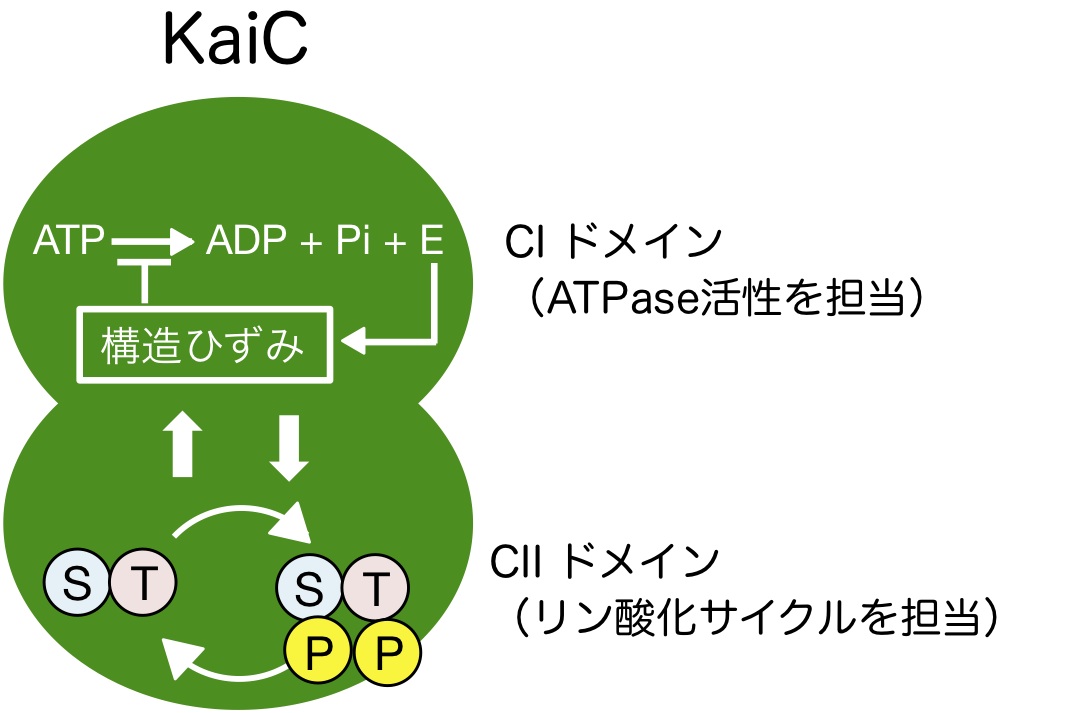
図7:CIのATPase活性によるエネルギーがKaiC自身の活性を抑制し、CIIのリン酸化サイクルと関わる。
執筆: H.I.& K.M.
 お問い合わせはこちらへ kondo@bio.nagoya-u.ac.jp
お問い合わせはこちらへ kondo@bio.nagoya-u.ac.jp〒464-8602 名古屋市千種区不老町 名古屋大学大学院 理学研究科 生命理学専攻 植物第一講座
TEL/FAX: 052-789-2498または2495